The Mystery of Proprioceptive Awareness
Published: February 24, 2022 | Updated: March 2026 | Author: Asia Shcherbakova
Although we are obviously consciously aware of the position of our body parts in space, it is often assumed that the signals from our muscles and joints that provide the input for the brain to calculate positional information do not reach our conscious awareness. We can feel touch on our skin—yes. But apparently, according to mainstream neuroscience, we cannot feel “tactile” sensations in our muscles (apart from pain and tension). But is it really true? Have you really never felt any neutral or pleasant sensations in your muscles? If the honest answer is “no, I don’t think so”—that turns out to be an equally interesting response, and this article is for you too. This article explores the topic of “proprioceptive awareness” as a dimension of body awareness, its possible neural substrate, and why—despite being a real and developable perceptual capacity—it has no established name in the scientific literature.
Introduction
Section titled “Introduction”If I were asked to make a list of benefits of physical exercise in order of importance, developing perceptual skills would come first—the ability to be constantly aware of the sensations in the muscles and of the positions of multiple points on the surface and inside the body. Any other benefits—being in shape, being healthy, feeling great, being productive—seem to naturally follow from this.
Yet my #1 benefit rarely makes it onto any such lists. Any catalogue of what exercise does for you will list health outcomes before it lists perceptual ones—if it mentions perceptual outcomes at all.
A commonly recognized category that overlaps with my #1 is “body awareness.” There’s quite a bit written about “the benefits of body awareness.” But the term is used inconsistently, and a clear definition is rarely provided. It may refer to a range of things—from kinesthesis to Craig’s interoception, or even to something close to Ellen Langer’s mindfulness.
My #1 refers to neither of these things. In our framework, it refers to a combination of what we call “proprioceptive awareness” and “spatial awareness.” This article will focus only on proprioceptive awareness. Because no common definition of “body awareness” exists yet, I’d like to first introduce our own framework for talking about it.
Body awareness is a complex, multi-dimensional construct in need of more nuanced conceptualization. A clear definition is rarely provided.
— Mehling et al., 2009
The 3 Types of Body Awareness
Section titled “The 3 Types of Body Awareness”In Baseworks, from a practical and pedagogical standpoint, we find it useful to distinguish among three types or dimensions of body awareness:
- Interoceptive awareness
- Proprioceptive awareness
- Spatial awareness
Interoceptive awareness is the conscious awareness of sensations related to internal organ functioning, stress, emotions, and metabolic changes. Our notion of interoceptive awareness is mostly in agreement with Price & Hooven’s interoceptive awareness as a perceptual skill extension of Craig’s idea of interoception.
The following two types are more distinct, and it would be difficult to match them precisely with a pre-existing framework or category.
Spatial awareness, in our framework, refers at the most basic level to the ability to feel the position of body parts in space without visual input (kinesthetic localization). Beyond this, in our framework, spatial awareness also relates to the ability to use a mental model of space to map it onto the external environment for various purposes—mnemonic, aesthetic, exploratory, recreational, or otherwise. It is related to my #1, but I won’t discuss it in this article.
Proprioceptive awareness, in our framework, is the conscious awareness of signals from the mechanoreceptors of the musculoskeletal system (as opposed to the skin), experienced as localized sensations.
Note: tactile sensations in the skin belong to the domain of “exteroception” (sensing the external environment) and are not covered by the 3 types of body awareness framework.
This classification is very much application- and task-based, and sometimes it’s unclear how to classify a particular sensation.
For example, when I am aware of my heartbeat—as in, having a pulsing sensation somewhere in my body without touching the skin with my fingertips—is it interoceptive or exteroceptive awareness? Although large blood vessels are equipped with specialized stretch-sensitive receptors, according to Wilfrid Jänig (my go-to source when it comes to the autonomic nervous system), their stimulation does not seem to give rise to conscious sensations. So perhaps the sensation of the heartbeat is actually picked up by the skin (maybe we could say that our exteroceptive capacity contributes to interoceptive awareness).
Or, if I am aware of the peristalsis in my transverse colon moving against my abdominal muscles (you know, the slow rhythmical contractions of the part of the large intestine that lies just above the belly button—coincidentally, or not so coincidentally, the spot where I used to get my first needle whenever I visited an acupuncturist in Tokyo), should it be considered interoceptive or proprioceptive awareness?
Although we don’t always have a clear answer, this framework still serves the needs of our teaching applications and research. It also helps solve two problems that tend to come up whenever “body awareness” is discussed:
- Mixing unrelated phenomena together (example: “body awareness is the awareness of the body position in space, but it also includes our awareness of being hungry”)
- Failing to notice certain phenomena because they ambiguously fall on the overlap between existing categories (example: “proprioception occurs beyond our awareness”)
Proprioception vs Proprioceptive Awareness
Section titled “Proprioception vs Proprioceptive Awareness”Before diving deep into the topic, I’d like to bring attention to one important terminological distinction.
Proprioception and proprioceptive awareness are not the same thing, even though they are frequently used interchangeably.
Proprioception is the system—the receptors, pathways, and brain circuits—that continuously provides the nervous system with information about muscle length, force, and joint position. Much of this happens without us noticing it, and it’s indispensable: without it, you couldn’t coordinate movement, maintain balance, or reach for a glass without looking at your hand.
Proprioceptive awareness, as we use the term, is something different. It is the conscious access to sensations produced by this system, experienced as localizable feelings within the muscles and joints.
This distinction between a perceptual system and the conscious awareness of its signals is not novel. The interoception literature makes a similar move: deep interoceptive signals—heart rate, gut motility, visceral states—operate largely without our awareness, but interoceptive awareness, the ability to consciously attend to and represent those signals, appears to be a trainable skill (Price & Hooven, 2018). As I will argue below, something analogous may be true for proprioception.
Proprioceptive Awareness, Really?
Section titled “Proprioceptive Awareness, Really?”The term “proprioception” has a wide range of meanings. Neuroscientists, for example, may treat somatic proprioception as an entirely subpersonal, nonconscious function.
— Shaun Gallagher (philosopher known for his work on embodied cognition, social cognition, agency, and the philosophy of psychopathology)
I know—“proprioceptive awareness” defined as conscious sensations may raise quite a few eyebrows. Can the proprioceptive system give rise to conscious, localizable sensations at all?
When your finger hurts, or someone touches it, you can locate the sensation in the finger and not in your neck or your stomach. That is what I mean by localizable, and this is a commonly shared experience. But textbooks say proprioception doesn’t work like that—that it operates below the threshold of localizable conscious sensation.
A significant strand of the scientific literature on ‘body awareness’ has focused primarily on interoceptive awareness. A 2009 systematic review of body awareness assessment tools found that the proprioceptive dimension appears only as a proxy for psychological distress—“sensations of distress, worry, pain and tension” (e.g., “I am aware of tension in my muscles”)—not as a domain of neutral or positive sensation in its own right (Mehling et al., 2009).
And yet:
Have you really never felt any sensations in your muscles beyond pain and soreness?
Take two identical glasses. Fill one with water at room temperature. Hold both in the air, one in each hand, and close your eyes. You can immediately tell which one is heavier. But if you pay careful attention to the sensations in your arms while doing this, you may also notice something else: a sense of more of something in the arm holding the heavier glass—something fuzzy, diffuse, but localizable within the muscle.
This is the “mysterious” sensation this article is about.
At this point I am anticipating that the readers’ perception will split. Some will wonder, confused, “What’s so mysterious about this sensation? Isn’t it just a normal human experience?” and some will wonder, also confused, “What is she even talking about?” I will address both these potential reactions.
In our own ongoing survey of non-practitioners (n=48 to date), roughly 47% of respondents report being aware of muscular sensations at rest, roughly 47% report awareness only during exercise, and the remaining 6% report not feeling any localized sensation in muscles even during physical activity. On top of this, nearly half of respondents familiar with the sensation at rest—48%—didn’t know what to call this sensation. It is as if sighted people had no vocabulary to talk about light or color.
| Not aware | Aware during exercise | Aware at rest |
|---|---|---|
| 6.4% | 46.8% | 46.8% |

Based on our survey data and my conversations with various people, the word that’s most commonly used for this sensation is activation. It is a conscious sensation localized within a muscle that either appears or intensifies when the muscle is contracted. Some people only experience it in a contracted muscle, not at rest. It’s immediate—not the feeling that builds with exhaustion. It’s not pain or tension. It’s not unpleasant. It feels like the normal quality of an engaged muscle, rather than anything that would make you stop.
The scientific literature, as far as I can tell, has no name for this sensation.
The philosophical literature, interestingly, hasn’t closed this gap either. Phenomenologists working in the tradition of Merleau-Ponty have taken proprioception seriously as a dimension of lived experience—exploring what it’s like to have a felt sense of the body in space: posture as orientation, movement as intentional gesture, the body as a unified presence. Gallagher’s own work on body schema and body image belongs to this tradition. But it addresses proprioception at a different level of processing and with a different focus. “Proprioceptive phenomenology,” as that tradition uses the term, tends to mean the spatial and kinesthetic experience of moving through an environment (which is more related to the perceptual domain of what we call spatial awareness)—not the texture of sensation inside a particular muscle. The more specific question—what does the muscle itself feel like from inside, without pain or post-deadlift lactic acidosis?—rarely surfaces there. It isn’t that the philosophical literature claims no such sensations exist. It simply has a different angle of inquiry, and the question doesn’t arise from within it.
The main purpose of this article is to bring attention to conscious sensations originating from the musculoskeletal system—primarily in the muscles—and to explore why this phenomenon has received almost no attention in scientific literature, despite appearing to be a real and developable perceptual capacity.
The following diagram summarizes what this article is about: this black hole, the blind spot, the void in an otherwise very regular system of how we tend to think about interoception, proprioception, and exteroception.
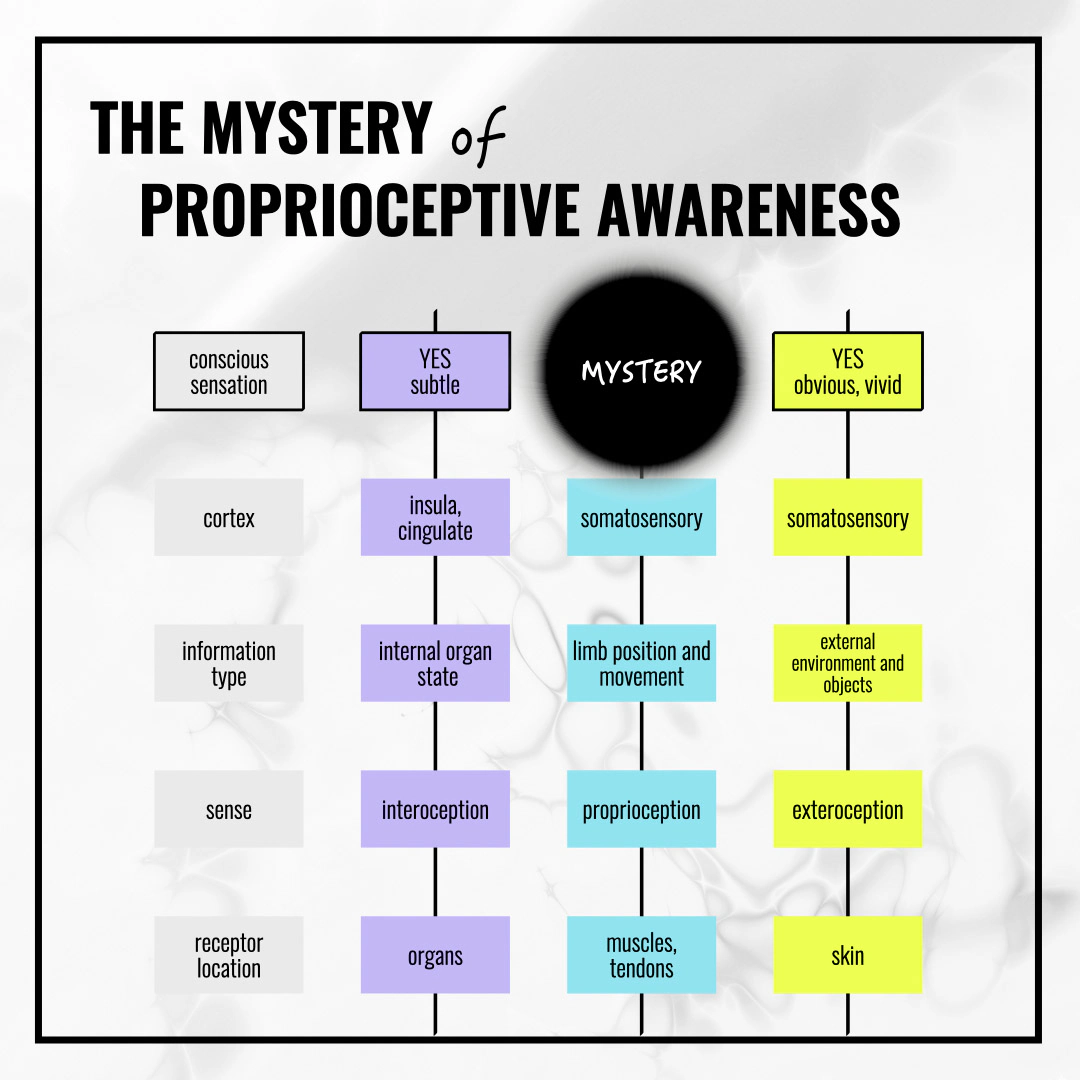
The blind spot: interoception and exteroception both produce recognizable conscious sensations. Proprioception follows the same route to the same cortex—but its phenomenology remains unexplored.
Why Is It a Mystery? The Three Dichotomies
Section titled “Why Is It a Mystery? The Three Dichotomies”Almost half of respondents in our survey reported being able to perceive localized sensations in their muscles even without active contraction. While our sample could be skewed towards more vivid muscle awareness because many people found the survey landing on the page from web search of body awareness-related topics, the trend still indicates that this is a wide-spread perceptual profile. Many people perceive these sensations and recognize their descriptions.
Yet, if you go looking for “conscious sensations in active muscles” in the scientific literature—not pain, not soreness, but neutral and localizable sensations—you’ll find almost nothing. There are, I think, three structural reasons why this topic falls through the cracks.
Dichotomy 1: “Conscious” skin sensations versus “unconscious” proprioception
Section titled “Dichotomy 1: “Conscious” skin sensations versus “unconscious” proprioception”The standard account contrasts touch—which everybody agrees is conscious and localizable—with proprioception, which is typically described as operating without awareness. According to this account, skin receptors feed the brain information that gives rise to felt sensation. Muscle receptors feed the brain information that gives rise to motor commands and position sense—but not, apparently, to felt sensations. So, we have a dichotomy between exteroception and proprioception.
This picture is not wrong. It’s simply incomplete. The fact that proprioceptive signals get converted by the brain into positional coordinates unconsciously doesn’t mean that no signal from the musculoskeletal system ever reaches conscious experience. But by consistently pairing “skin” with “conscious sensation” and “muscle and joint receptors” with “unconscious proprioception,” the standard framework leaves no conceptual place for sensations in muscles that are neither pain nor soreness.
A good example of this view is this review (Delhaye et al., 2018). https://onlinelibrary.wiley.com/doi/abs/10.1002/j.2040-4603.2018.tb00045.x
Dichotomy 2: “Conscious” proprioception versus “unconscious” proprioception
Section titled “Dichotomy 2: “Conscious” proprioception versus “unconscious” proprioception”Even within proprioception itself, there is a split. According to this account, the dorsal-column medial lemniscus (DCML) pathway carries somatosensory information from muscles and joints up to the cortex, where it contributes to conscious positional sense—the ability to tell where your arm is without looking. A separate proprioceptive stream feeds into the cerebellum, where these signals are used for continuous real-time motor coordination with no accompanying awareness.
So, sometimes, when people talk of “conscious” and “unconscious” proprioception, they draw the distinction between the cortical and cerebellar (subcortical) streams. The cerebellar processing is considered truly unconscious, while the DCML pathway that feeds into the positional sense is identified as the “conscious” aspect of proprioception. Again, this is not wrong, but leaves no room for localizable muscular sentience.
A good example of this view is this recent book chapter, especially Figure 2 (Vega and Cobo, 2021), and many popular educational sources, such as The Ascending Tracts page on TeachMeAnatomy. https://teachmeanatomy.info/neuroanatomy/pathways/ascending-tracts-sensory/
Dichotomy 3: The sensory-motor homunculus split
Section titled “Dichotomy 3: The sensory-motor homunculus split”Most people interested in the topic of body awareness have encountered the cartoons of the sensory homunculus and the motor homunculus (which look like a deformed person laid over a strip of brain cortex) – the graphic representation of the sensory and a motor maps discovered by a Canadian neurosurgeon Wilder Penfield by poking brains of his epileptic patients with electrodes during open-brain surgery. The explanation almost always uses touch as the reference: “if someone touches your finger, you know where you’ve been touched because of this map.” The motor homunculus, adjacent to it, represents muscles as outputs—the “motor” side.
This creates a powerful implicit framework: sensory = input = skin, motor = output = muscles. The possibility that muscles might also produce sensations that could, in principle, reach conscious awareness is not explicitly denied, but it is implicitly suppressed because this framing reinforces the input-output distinction.
There is no single source for this because the assumption is rarely stated explicitly. Instead, it is embedded in how the topic is typically presented across neuroscience textbooks and educational resources.
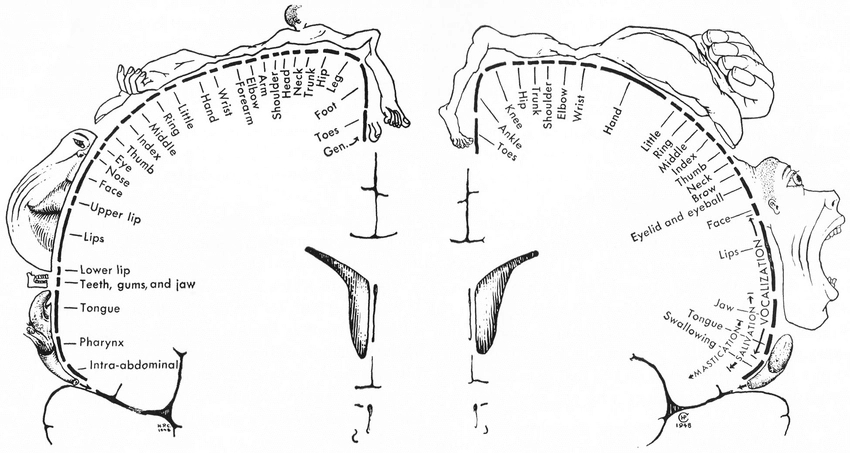
Penfield’s homunculi (1948). The sensory map is almost always explained using touch: “if someone touches your finger, you know where, because of this map.” The motor map represents muscles as outputs. The possibility that area 3a—adjacent to the skin-sensation areas, receiving muscle and joint input—might also produce localizable sensation is not part of this framing.
These three dichotomies reinforce each other. Together, they create a structural blind spot: neither framework denies this explicitly, but collectively, they leave no room for conscious, localizable sensation in muscles. This sensation doesn’t belong to any existing category—not tactile, not pain, not positional sense, not unconscious proprioception—so it tends not to be studied, described, or named.
Where Are the Muscle-Associated Sensations in the Brain?
Section titled “Where Are the Muscle-Associated Sensations in the Brain?”Someone who has never paid attention to muscular sensations might reasonably ask: “Why should there be conscious, localizable muscle sensations in the first place?”
The answer begins with how sensory signals travel from the body to the brain. The nervous system uses dedicated pathways—specific cables, each carrying a particular type of information from particular receptors to a dedicated region in the cortex. The pathway most relevant here is the dorsal column–medial lemniscus pathway, or DCML.
The DCML carries signals from two distinct sources in close parallel: mechanoreceptors in the skin (responding to pressure, vibration, and texture) and mechanoreceptors in the muscles and joints (responding to stretch, force, and joint position). Both travel directly to the primary somatosensory cortex—three synapses from receptor to cortex. The same number for skin. The same number for muscle.
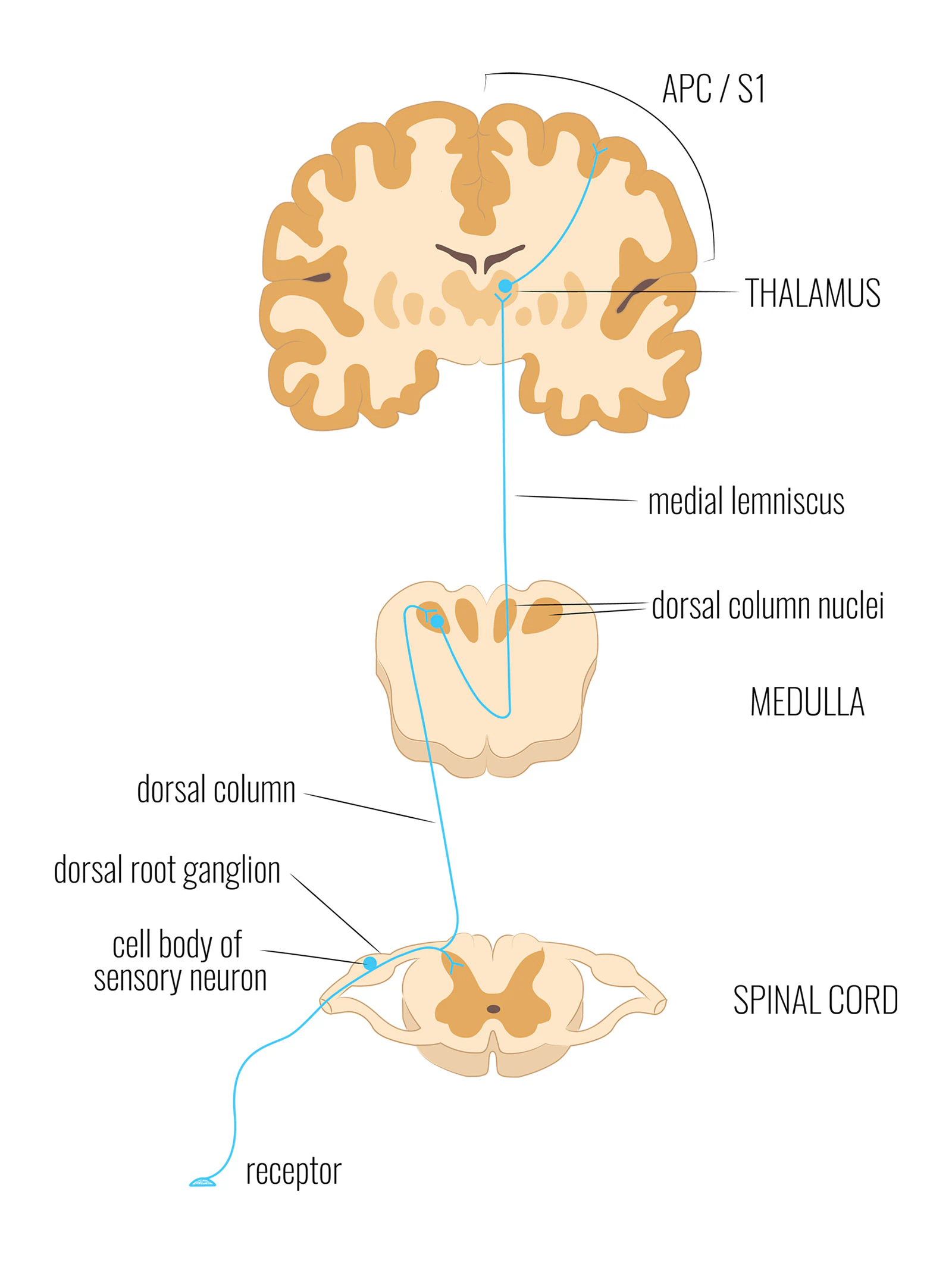
The DCML pathway: from receptor to cortex in three synapses. Both skin mechanoreceptors (pressure, vibration, texture) and muscle mechanoreceptors (stretch, force) travel this same route to adjacent areas of the same cortex.
For touch, this is exactly how we explain conscious localization: skin signals arrive in S1 region that corresponds to a specific location in the body and give rise to the felt sensation of being touched at a specific location on the body. It is commonly said that S1 contains “a map” of our skin surface.
The primary somatosensory cortex is not a single uniform area. What is commonly called “S1”—more precisely, the anterior parietal cortex—consists of four parallel strips: areas 3a, 3b, 1, and 2.
- Areas 3b and 1 primarily receive information from the skin
- Area 3a receives primarily proprioceptive input—from muscles and joints
- Area 2 receives input from both
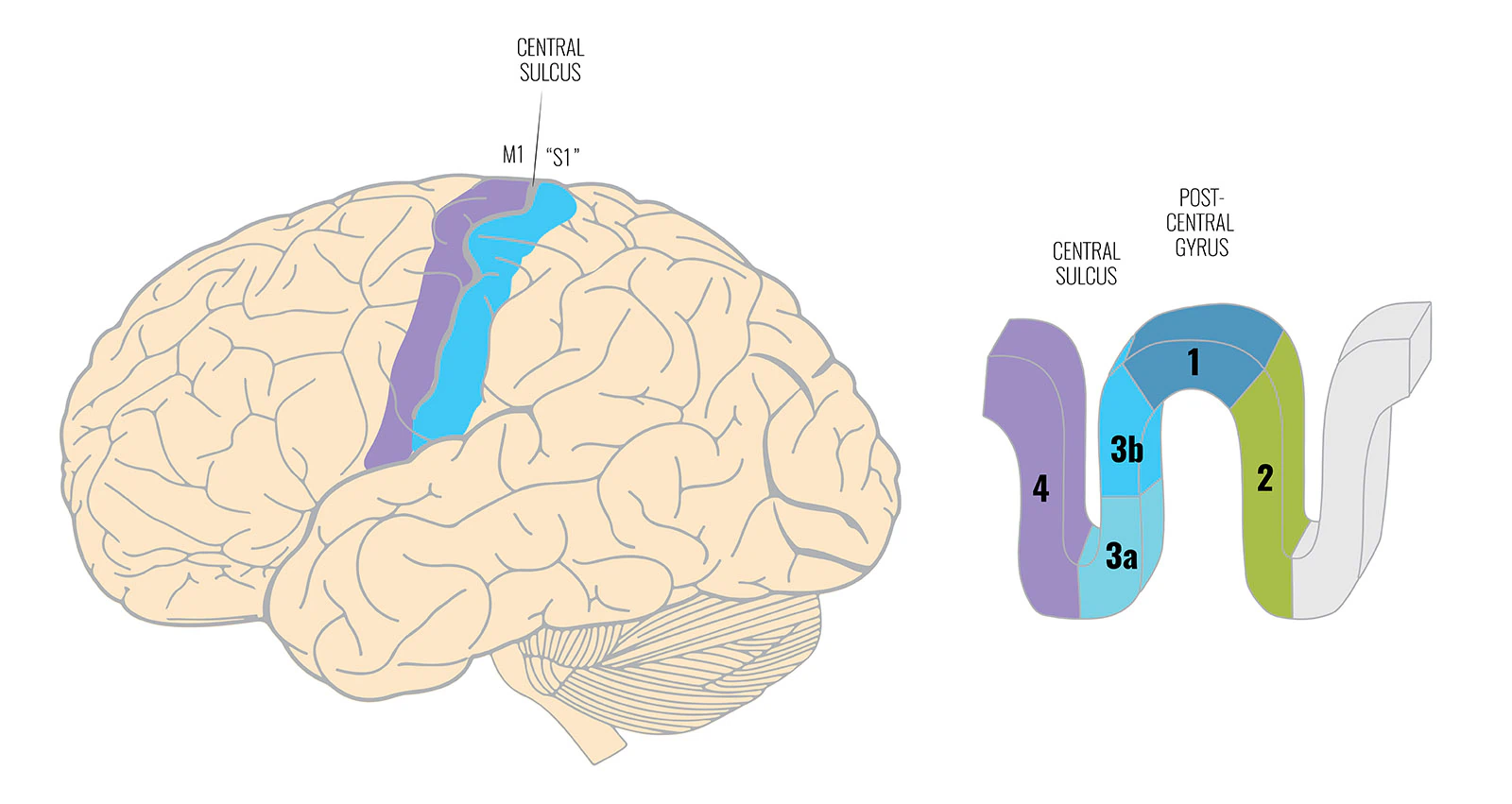
S1 is not a single area. The anterior parietal cortex comprises four parallel strips: areas 3b and 1 receive skin input, area 2 receives mixed input, and area 3a—the deepest, closest to the motor cortex—receives proprioceptive input from muscles and joints.
The standard explanation of the sensory homunculus uses touch: if someone touches your hand, you know where because this map localizes the signal. Areas 3b and 1 are the substrate for that conscious localization. Immediately adjacent, area 3a receives the muscle and joint signals.
However, here the trail goes cold.
Skin signals and muscle signals travel the same route, arrive at adjacent areas of the same cortex. One set unambiguously gives rise to conscious, localizable sensation. The other, the standard account says, does not—it is converted into positional coordinates without producing anything you might feel. The question of what a healthy, pain-free person actually experiences from area 3a—if anything—appears not to have been systematically asked.
One of the leading researchers in proprioception confirmed this gap in personal correspondence. He reported that he himself does not experience sensations in relaxed muscles, and noted that he was not aware of any literature on the topic of constant conscious muscular sensations in healthy subjects (personal communication). He added: “You are right it is 3a which is the primary receiving zone for muscle afferents, but I have not ever read a report that stimulating 3a generates sensations in passive muscles.”
Area 3a remains the most logical candidate for where such sensations would be processed. But the experiment that would close the question appears, on this account, not to have been done.
What Is This Sensation?
Section titled “What Is This Sensation?”So, what exactly is this sensation? Why do some people feel it and some don’t? What is its function? Should you even care about it? Can you find it if you don’t have it?
What I describe here is a hypothesis which is based on a decade-long survey of scientific literature, pedagogical experience, and phenomenological discussions with different people.
If there is a conscious sensation, there must be a receptor. The most abundant type of receptor in the muscle is the muscle spindle, often called the stretch receptor. They sit between force-producing muscle fibers, and when a muscle lengthens, they increase their firing rate. This signal is sent to the brain to signal the length of the muscle and the speed of length change, and the brain uses this information to compute joint angles and how the body moves in space. As stretch receptors, they respond to muscle lengthening and, at first glance, seem like a poor candidate for a sensation that intensifies with muscular contraction.
Skeletal muscle also has some pressure receptors (Paciniform receptors, Ruffini type receptors), which are similar to the cutaneous (skin) receptors, only much less abundant. And to be honest, when I first started thinking about this a few years ago, my first instinct was that this sensation was associated with these pressure receptors.
But long story short, the spindle story is more complex than it first appears. In fact, it’s somewhat misleading to think about a spindle as “a receptor.” When we think about receptors in biology, we might conceptualize them as specialized cells, signaling a particular modality, where the firing rate varies but the signal type is fixed. Perhaps like rods and cones in the eye. But the spindle isn’t like that. It is more correct to think about a spindle as a sensory organ containing several receptors. It’s a capsule that contains two distinct types of sensory endings with genuinely different jobs: Group Ia and Group II fibers that carry different kinds of information to the brain.
(spindle illustration: extrafusal and intrafusal fibers, alpha motor, gamma motor, group 1 afferent, group 2 afferent fibers)

Group Ia fibers. They are attached to the central part of the fibers inside the spindle. These are velocity-sensitive and phasic. They fire hard when the muscle is suddenly stretched and quickly go quiet. This is the signal behind the stretch reflex, and it’s the dominant reason spindles are called stretch receptors.
Group II fibers. These are attached at the end part of the fibers inside the spindle. These fire continuously, proportional to how taut the spindle is right now—regardless of whether the muscle is being actively stretched or has been still for some time (Vallbo, 1974). This signal doesn’t go to zero at rest. It reflects the current mechanical state of the spindle. It requires no motor command (ongoing action) to maintain. This is the most plausible substrate for the resting baseline sensation in the muscle.
Why does the brain need to know “how taut spindles are”? This is where it gets even more interesting and why the phenomenology of spindle sensations is so under-discussed.
When the muscle shortens, the spindle would normally go slack—like a guitar string whose neck has been shortened. A loose string produces no sound. A slack spindle can’t signal. To be sensitive to stretch (its main function), the spindle has to be taut.
The neuromuscular system solves this with a dedicated mechanism. Alongside the motor neurons that tell the main force-producing muscle fibers to contract (alpha motor neurons), a separate set of neurons (gamma motor neurons, also called fusimotor neurons) contracts the ends of the spindle, keeping it taut in the middle as the surrounding muscle shortens. Alpha motor neurons and gamma motor neurons are co-activated together. This is called alpha-gamma coactivation. The effect: a spindle that stays taut keeps firing—both the Group II tonic signal and the Group Ia signal are intensified, proportionally to the level of contraction (with some desensitization at high force, as I will mention below). The ascending signal generated by this mechanism is what researchers call fusimotor reafference. (“afference” means “signal going to the brain”, and “reafference” means that it is a signal that goes “back” to the brain in response to the motor signal generated by the brain).
So the group II secondary afferents communicate to the brain “how taut this spindle is,” not “how much the muscle where this spindle lives is stretched.” So, it is somewhat of a housekeeping signal that perhaps doesn’t require much conscious awareness or attention. Just think about it: when was the last time you had to make a decision based on how taut the spindles in your muscles were? If you never needed this information, there is a high chance that you just never paid attention to it.
If you were to google diagrams of muscle spindles, you’ll notice that many of them only feature the alpha motor neuron (that contracts the force-producing muscle fibers) and the Group Ia afferents. This reflects how the spindle story is usually told—it is a stretch receptor that fires when the muscle lengthens. Some diagrams would also feature gamma motor fibers but not Group II fibers—again, telling the story of the “stretch receptor” that needs alpha-gamma co-activation to continue functioning as a “stretch receptor” when the muscle shortens. Of course, some diagrams properly include Group II afferents, as well as other parts that I didn’t name. Still, the trend we can see across all the diagrams represented across the internet is the secondary, supporting role of the Group II afferent fibers while the Group Ia fibers take the stage. I don’t think I’ve ever seen a diagram of photoreceptors in the eye that forgot to include the sensory fibers that carry the visual signals to the brain. So, the sensory story of the Group II fibers is being systematically silenced by the dominant “stretch receptor” narrative. I am being a little over-dramatic here on purpose, but I do find it a little bit sad that the Group II “tautness” signal has been measured for fifty years yet the phenomenology was never the question anyone thought to ask.
And this brings me to an important paper that comes closest to the perceptual correlates of Group II afferent inflow in a contracting muscle.
What Luu and colleagues showed in 2011 is that fusimotor reafference (the sensory signals that spindles send to the brain when a muscle is contracted) is a major contributor to our sense of force and heaviness when we lift objects. This was a novel finding because it was previously assumed that the sense of effort is generated internally by the brain. The researchers in this study reduced spindle sensitivity in subjects by using high-force fatiguing contractions (which desensitize the spindle receptors), then had them lift a weight. The weight felt lighter—not heavier, as you’d expect if the brain’s internal effort signal were doing the work. They also measured this in subjects with a rare neurological condition that eliminates spindle feedback entirely—the opposite happened: objects felt twice as heavy after fatigue. The spindle signal, not the central effort estimate, appears to be the dominant contributor to force perception in healthy subjects.
So while Luu and colleagues did not study the phenomenon of localized muscular sensations, they demonstrated that the conscious felt sense of weight and effort in a contracting muscle comes from the spindles—the stretch receptors.
So, hypothesizing the neural basis of the localized muscular sensations, there are likely two components:
- a tonic baseline that is always present when the muscle is at any length, coming from Group II fibers and
- a contraction-proportional intensification driven by fusimotor co-activation.
Analyzing the phenomenology of this sensation—specifically, its continuity between rest and contraction—I’ve found myself reaching for the word hum. Because it is like a hum: present at a low register when the muscle is at rest and intensifying as contraction increases. The same phenomenological stuff, going from quiet to loud.
In Baseworks, we use the word activation when directing attention to these sensations in active tasks—specifically the contraction-proportional intensification, where the signal rises and becomes more legible. But “activation” captures only that louder part. “Hum” is the word I use for the full phenomenon, including its resting baseline.
This is explicitly a hypothesis. Luu’s paper addresses force and weight perception, not localized muscular sensation as a distinct subjective experience. The step from “spindle reafference contributes to force perception” to “the hum is spindle Group II afference reaching conscious awareness” is logical but not experimentally confirmed. The researcher I mentioned earlier said it plainly: no report of stimulating area 3a producing localizable sensations in passive muscles appears to exist. The question is open.
Another word that comes close to naming this in practice is the word “tone.” The clinical term “muscle tone” refers to the low-level resting tension maintained in muscle tissue. However, some practitioners with extensive movement training use this word for the sensation itself—not for what a clinician measures, but for what they feel. Matan Levkovich, a dancer and movement educator at Movement Lab [link to movement lab], describes it using “muscle tone” for exactly this sensation—a term he had adopted from someone in his own training. The guitar string analogy is useful here too: tone in music refers to a string under enough tension to vibrate. What the fusimotor system maintains—keeping the spindle sensitive during contraction—is something like the sensory equivalent of that.
”If I Cannot Feel Anything, I Am Not Doing Anything”
Section titled “”If I Cannot Feel Anything, I Am Not Doing Anything””There’s a widespread movement and exercise mindset: more is better. If you look at how exercise is culturally promoted—the language used, what counts as a productive session, what kind of sensation is presented as the sign that something real is happening—there’s a persistent emphasis on strong stimulation. The burn. The pump. The feeling of effort. The endorphin release. The emphasis on weight training as the most effective path to longevity.
There’s a lot I could say here about why this narrative of overstimulation might be counterproductive long term for a lot of people, but this falls outside the scope of this article. What I’d like to focus on here is how this narrative becomes yet another factor that keeps muscular sensations hidden, adding to the mystery.
Spindle primary afferents (group Ia) progressively desensitize during sustained high-force contraction—their firing rate roughly halves within a minute of sustained effort at 20% of maximum voluntary contraction, and the desensitization is greater at higher force levels (Macefield et al., 1991). Luu’s experiments showed this directly: after high-force fatiguing contractions, subjects perceived lifted weights as lighter, not heavier. The spindle signal had been blunted. So the very intensity that the exercise narrative associates with a meaningful workout actively degrades the signal that may underlie the sensation of activation.
There’s a second mechanism that compounds this. During intense exercise, many other signals are competing for cortical attention: the burn from lactic acid (carried by small-diameter group III and IV afferents), cardiovascular arousal, the overwhelming sense of effort. These signals are loud. You may have noticed that rubbing a sore spot reduces the pain while you’re rubbing it—this is a related phenomenon, where strong input in one channel suppresses the experience of input in another. When these signals are running loud, the subtle, localized fusimotor reafference gets selected against—not because it isn’t there, but because attention has nowhere left for it.
There is also a third mechanism that may be the most fundamental. Perception requires calibration: to locate a signal at a lower intensity, you need to have attended to it at a higher intensity first. If you have never experienced a clear, legible instance of the spindle hum—never attended to it during deliberate moderate contraction where it is salient enough to anchor in experience—you have no perceptual reference threshold from which to locate it at rest. The resting signal is there—and your brain continues to use it in the background in its computations to deliver the “conscious” positional sense —but without a calibrated reference it remains in the background, beyond the threshold of your conscious awareness. This is part of why practices that maintain intensity just high enough to make the hum legible—while keeping it well below the level that desensitizes the spindles or generates competing sensory noise—create conditions where calibration actually becomes possible.
The full chain looks something like this: cultural narratives push toward high intensity → high intensity desensitizes the spindle receptors directly, while simultaneously flooding the sensory system with competing signals → the perceptual system never calibrates to the hum → the sensation never becomes a salient, attended perceptual object → nobody develops vocabulary for it → because there is no vocabulary, there is no discussion → because there is no discussion, the science doesn’t study it → the sensation has no name.
And at the opposite end—various “mindfulness” practices that cultivate stillness, or very slow, unfocused movement without deliberate contraction—don’t produce the contraction-proportional intensification of the hum.
What develops conscious access to that baseline signal is a different technique: creating the contraction-proportional intensification, and then deliberately lowering it as far as possible, so the resting signal can be calibrated against what has just been felt. Stillness alone never creates that rise and return to baseline. Vigorous dynamic movement tends to generate rapid phasic changes that overwrite the subtle signal rather than isolate it. Neither, on its own, builds the perceptual reference needed to attend to what’s there at rest.
Because perception is, at its core, a skill that requires attention to salient details: if the attention to low-intensity muscular sensations has never been systematically cultivated, and if every cultural narrative about movement points toward stronger stimulation as the sign of real work, it’s not surprising that this particular signal has never become a recognized perceptual category.
What’s the Usefulness of Proprioceptive Awareness?
Section titled “What’s the Usefulness of Proprioceptive Awareness?”I personally didn’t pay any attention to these sensations until I was almost 30. And it was my involvement with Baseworks and one of its key techniques—Distributed Activation, which requires simultaneous low-level co-contraction of every muscle in any movement—that started to shift my sensory threshold. What had been barely noticeable became gradually, and then quite substantially, part of my ongoing experience.
Currently, the sensations coming from the muscles and the sense of position in space constitute a large proportion of my moment-to-moment sensory experience. They are—and I realize this might sound strange—genuinely entertaining and rewarding. Of course, sometimes I have doubts: maybe this is something that happens naturally in one’s 30s and has nothing to do with the practice. But I don’t remember my grandmother, or my mother, or anyone else teaching me that at 30 one suddenly begins to find the sensation of simply sitting upright fascinating. And when I try to describe these experiences to other people, there’s often no shared vocabulary and no reference to a shared experience. To the point that when I read about “dopamine hits” from scrolling through social media, I genuinely wonder: really? How can that compete with the intrinsically rewarding sensation of being aware of the trajectories of different body parts when walking?
Which is a long way of saying: it just feels very nice, and it’s available all the time.
Beyond the intrinsic quality of the experience, proprioceptive awareness has direct practical relevance in movement learning. It’s more than a perceptual curiosity because the relationship between sensory access and motor control is tightly interconnected.
Being able to feel a muscle makes it easier to control that muscle. Moreover, it seems like training the ability to control a muscle in isolation often comes with developing a dedicated feedback system for that particular muscle. In teaching practice, we hear reports from students on how muscle sensations “pop up” from silence and “split,” correlating with one’s ability to control muscles in isolation, which could be explained by cortical remapping following functional separation, similar to what has been reported after surgical separation of fused fingers in syndactyly (Mogilner et al., 1993). The mechanism is not unusual, but the phenomenology is. When someone says, “I couldn’t feel anything here and now I can suddenly feel it,” this typically comes with awe and surprise. This isn’t easy to track as a training outcome—unlike movement range or force output, there’s no simple metric or predictable timeline for it—but the reports we hear in teaching are consistent.
When we teach Distributed Activation (co-contraction of as many muscles as possible in any movement at low intensity), inexperienced practitioners often report not being able to determine “how much” to contract the muscles. I find it fascinating because from my sensory perspective, the answer is simple: just pay attention to the hum and create a uniform sheath of it across all your muscles. But of course, this is impossible if you don’t perceive it in every muscle and if it’s uncalibrated. While it’s possible to control a movement without feeling strong sensations in a muscle, training to calibrate proprioceptive awareness generally correlates with the ability to control individual muscles.
In practice, we see different patterns of how low proprioceptive awareness manifests, especially when it comes to learning the Distributed Activation. Some people tend to underactivate, moving loosely without co-contraction. Others tend to hyperactivate, as if they were doing high-intensity isometric holds as part of a strength-training routine. In both cases, what we would infer is a perceptual gap: without an accessible felt reference for what the right level of activation is, ‘more’ or ‘less’ doesn’t connect to anything the practitioner can actually identify and control.
How to Develop Proprioceptive Awareness
Section titled “How to Develop Proprioceptive Awareness”The general principle here is straightforward: if there is a receptor, and if its pathway reaches the cortex, conscious discrimination ability in that modality can be improved through practice—through the deliberate and repeated act of attending to the corresponding sensation. This applies to taste, to color discrimination, to sound. There’s no obvious reason why it wouldn’t apply to fusimotor reafference and the baseline spindle inflow.
But the conditions matter. To summarize once again what we know from muscle neurophysiology research, at high intensity, the relevant receptors desensitize, and competing signals crowd out the subtler fusimotor input. In stillness, the contraction-proportional component isn’t generated—the resting hum is present, but without the contrast of a deliberate rise and return, the perceptual system has no opportunity to calibrate to it. A body scan during meditation targets a different perceptual layer—interoception rather than the musculoskeletal sensations we’re discussing here. A body scan could capture the existing patches of hum, but it won’t raise the perceptual threshold across the muscles which are silent.
What seems to work is deliberate, low-to-moderate intensity activation with consistent attention directed toward the sensations it produces. In Baseworks, these conditions are consistently created through Distributed Activation and Micro Movements.
The framework for understanding why this works emerged from the practical work itself—from many years of observing what actually helped people develop access to these sensations in teaching. We call it Proprioceptive Awareness because that is the most precise term available, even though the scientific literature has not given this specific perceptual capacity its own name.
That gap—between what can be developed in practice and what science has a word for—is itself part of the mystery.
Muscle Activation Sensation Questionnaire
Section titled “Muscle Activation Sensation Questionnaire”When I published the first version of this article in 2022, I didn’t know that so many people did not experience localized muscular sensations at rest. This knowledge came from the data accumulated through the questionnaire below.
If you’d like to contribute your experience to our ongoing data collection, the form is below. It can be completed anonymously without entering your name and email.
(form goes here)
References
Section titled “References”Craig AD (2015). How Do You Feel? An Interoceptive Moment with Your Neurobiological Self. Princeton, NJ: Princeton University Press. 10.1515/9781400852727
Delhaye BP, Long KH, Bensmaia SJ. Neural basis of touch and proprioception in primate cortex. Comprehensive Physiology. 2018 Oct 17;8(4):1575–602.
Gallagher S. Bodily self-awareness and object perception. Theoria et Historia Scientiarum. 2007;7(1):53.
Luu BL, Day BL, Cole JD, Fitzpatrick RC. The fusimotor and reafferent origin of the sense of force and weight. J Physiol. 2011;589(13):3135–3147.
Macefield G, Hagbarth KE, Gorman R, Gandevia SC, Burke D. Decline in spindle support to alpha-motoneurones during sustained voluntary contractions. J Physiol. 1991;440:497–512.
Mehling WE, Gopisetty V, Daubenmier J, Price CJ, Hecht FM, Stewart A. Body awareness: construct and self-report measures. PLoS One. 2009 May 19;4(5):e5614.
Mehling WE, Wrubel J, Daubenmier JJ, et al. Body awareness: a phenomenological inquiry into the common ground of mind-body therapies. Philos Ethics Humanit Med. 2009;4:6.
Mogilner A, Grossman JA, Ribary U, Joliot M, Volkmann J, Rapaport D, Beasley RW, Llinas RR. Somatosensory cortical plasticity in adult humans revealed by magnetoencephalography. Proc Natl Acad Sci. 1993 Apr 15;90(8):3593–7.
Price CJ, Hooven C. Interoceptive awareness skills for emotion regulation: theory and approach of mindful awareness in body-oriented therapy (MABT). Front Psychol. 2018;9:798.
Tresilian J. Sensorimotor control and learning: An introduction to the behavioral neuroscience of action. Bloomsbury Publishing; 2012.
Vallbo AB. Afferent discharge from human muscle spindles in non‐contracting muscles. Steady state impulse frequency as a function of joint angle. Acta Physiol Scand. 1974 Feb;90(2):303–18.
Vega JA, Cobo J. Structural and biological basis for proprioception. In: Proprioception. IntechOpen; 2021.
Related
Section titled “Related”- science—current scientific framing, §1.7 (Fusimotor Reafference)
- Distributed Activation
- Micro Movements
- Proprioceptive Awareness
- 3 Types of Body Awareness